Da quanto detto finora risulta evidente che ogni singolo colore percepito può essere sia l'effetto di una radiazione monocromatica (ad esempio un'onda a banda ristretta di 700 nm in grado di generare la visione del rosso) sia l'effetto del sommarsi in un'unica stimolazione di più radiazioni, ciascuna di lunghezza d'onda differente. Questa osservazione porta in primo piano un'importante caratteristica della percezione visiva, che la rende profondamente differente, ad esempio, dalla percezione uditiva. Mentre, infatti, l'orecchio è in grado di discriminare, in un accordo musicale, le singole note componenti, l'occhio non è in grado di separare, in una stimolazione luminosa composta dalla mescolanza di più luci diverse, le singole frequenze componenti.
La percezione visiva è sintetica piuttosto che analitica: una luce rossa ed una luce verde che colpiscono insieme un medesimo punto della retina avranno come risultato la percezione del giallo; non vedremo né il rosso né il verde.
Ciò significa che, nel contatto della radiazione elettromagnetica con i recettori della retina, l'informazione sulla lunghezza d'onda si perde. Al suo posto rimane la misura dell'eccitazione suscitata, che è proporzionale sia all'intensità della luce incidente sia alla sensibilità del recettore in quella particolare zona dello spettro a cui appartiene la radiazione che lo ha colpito.
Possiamo chiederci a questo punto: basterebbe un solo tipo di coni per spiegare la possibilità innata nella nostra vista di discriminare nella luce, allo stesso tempo, intensità e colori differenti?
La risposta a questa domanda è no. Per capire il perché occorre considerare il grafico in fig.6. La curva mostrata nel grafico rappresenta il differente grado di sensibilità di un unico ipotetico tipo di recettori fotosensibili, rispetto a luci monocromatiche di differente lunghezza d'onda. Nell'esempio, la radiazione monocromatica A1 ha una lunghezza d'onda corrispondente quasi alla massima sensibilità del recettore. L'effetto sul sistema percettivo si traduce in un valore 75 (puramente indicativo) di stimolazione, corrispondente alla visione di un certo colore avente una data intensità luminosa. La radiazione monocromatica A2 ha la medesima intensità della radiazione A1, ma ha una lunghezza d'onda differente, alla quale il recettore risulta tre volte meno sensibile, per cui l'effetto è una stimolazione tre volte meno intensa.
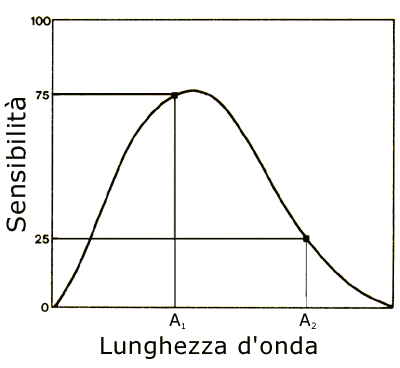
Fig. 6 – Due radiazioni
monocromatiche di differente lunghezza d'onda
in un sistema dotato di un unico tipo di recettori
Ora è dimostrato che in un sistema di questo tipo, in cui l'informazione sulla lunghezza d'onda si perde, lasciando in sua vece la percezione dell'intensità dello stimolo, è possibile eguagliare l'intensità della percezione dipendente dalla radiazione A2 all'intensità della percezione dipendente dalla radiazione A1 aumentando di tre volte l'intensità della radiazione A2. Ciò significherebbe, però, che la percezione visiva suscitata da A1 sarebbe del tutto uguale, indistinguibile da quella suscitata da A2, ovvero non ci sarebbe più discriminazione del colore, dal momento che vedremmo come uguali due radiazioni monocromatiche dotate invece di differente lunghezza d'onda.
In realtà questo è ciò che accade, all'incirca, nella visione scotopica dipendente dai soli bastoncelli: di tali recettori esiste infatti un solo tipo, che non è in grado, per i motivi appena descritti, di conservare l'informazione sulle differenze di lunghezza d'onda ma solo quella relativa alle variazioni di intensità luminosa.
Per avere allo stesso tempo discriminazione dell'intensità luminosa e del colore abbiamo bisogno di almeno due tipi differenti di recettori sensibili al colore. Con due tipi di recettori diventa possibile, ad esempio, eguagliare la percezione visiva dipendente da una singola radiazione monocromatica alla percezione visiva dipendente dalla miscela di due radiazioni monocromatiche di differente lunghezza d'onda.
Al principio dell''800 il fisico inglese Thomas Young propose una teoria della visione in cui si sosteneva la presenza di tre differenti tipi di recettori, ognuno dei quali in grado di percepire un particolare colore: dalla combinazione delle sensazioni provenienti da ciascuno di essi, risulterebbe la percezione dei colori nello spettro visibile. Nella sua ipotesi iniziale, Young indicò come colori primari - cioè quelli alla base di ogni possibile combinazione - il rosso, il giallo e il blu. Successivamente modificò la sua teoria indicando come primari il rosso, il verde e il violetto.
Le tesi di Young furono riprese circa mezzo secolo dopo dal tedesco austriaco Hermann von Helmholtz. Da allora la cosiddetta teoria tricromatica della visione, basata cioè sull'azione combinata di tre diversi tipi di recettori fotosensibili, è nota anche come teoria di Young-Helmoltz.
Si dovette aspettare ancora, però, circa un secolo, per avere – grazie alle rilevazioni effettuate nel 1964 con sofisticate tecniche di microspettrofotometria - la conferma sperimentale dell'esistenza di questi tre diversi tipi di recettori e delle loro specifiche sensibilità nei confronti della lunghezza d'onda della radiazione elettromagnetica. Il diagramma in fig. 7 illustra appunto le curve di sensibilità dei tre tipi di coni sperimentalmente individuati.
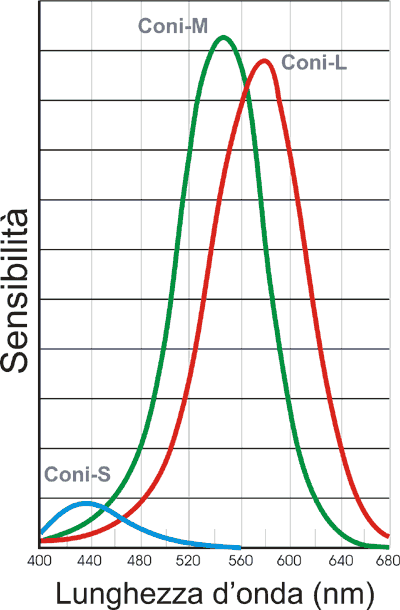
Fig. 7 – Curve di assorbimento della
luce da parte dei
tre tipi di coni sperimentalmente individuati
Le differenti posizioni, rispetto alla lunghezza d'onda, dei picchi di assorbimento della luce da parte dei tre tipi di coni dipende dalle differenti caratteristiche del pigmento – la iodopsina – in essi contenuto.
Dato un simile modello tricromatico di percezione dei colori, la visione, ad esempio, del colore giallo è l'effetto di una situazione in cui i coni-M (sensibili al verde) ed i coni-L (sensibili al rosso) sono massimamente stimolati, mentre l'eccitazione dei coni-S (sensibili al blu) è del tutto trascurabile. La visione del bianco si ha, invece, quanto tutti e tre i tipi di coni risultano massimamente stimolati.
Chi ha qualche esperienza dei modelli di rappresentazione del colore su computer avrà già capito che la teoria tricromatica della visione è l'antecedente fisiologico del modello di colore RGB.
![]() Leggi
La teoria dei processi opposti
Leggi
La teoria dei processi opposti
![]() Vai al
sommario
Vai al
sommario
![]() Scrivi a
info@diodati.org
Scrivi a
info@diodati.org
![]() Aggiornato
Aggiornato